Bilateral is a hypothetical organism, Urbilateria, and was the last common ancestor of Deuterostomia (echinoderms and the various chordate phyla) and Protostomia also referred to us higher animals. Classically, these two groups, the Deuterostomia, and Protostomia differed in a number of respects. Some of these characteristics have been challenged recently, but they remain generally true. One of the distinguishing factors is the cleavage pattern and the cells division in the early embryo. Protostomes are characterized by cell division running diagonally and vertically across the embryo. Each embryonic cell is established very early in the embryo before maturity (Palaeos.com 2002).
In Deuterostomes development, they undergo some radial changes that run parallel or perpendicular to the vertical axis of the embryo. They also obscure cleavage such that each early embryonic cell is independent enough to retain the capacity to develop into a complete embryo in case it’s isolated from other cells. During the first opening of the archenteron (blastopore), an event that occurs during gastrulation, unusual symmetry occurs. In Protostomes, the blastopore brings out the mouth first while in Deuterostomes the mouth comes out second and the blastopore forms the anus. The second difference is evident in the way the mesoderm surrounds the body cavity. It can either pod off or break apart. In Protostomia for example, the mesoderm also known as archenteron, splits part while in Deuterostomia, the mesoderm podes off (Palaeos.com 2002).
Biologically, the first “animals” were characterized by hollow balls of cells closely related to the living choanoflagellate colonies, dissimilar to the Fungi classifications and the clusters were clonal meaning they all evolved from a single parent cell. This characteristic qualifies them for a single organism classification. The first animals became experts at manipulating cell-cell interactions since it was difficult to maintain a hollow ball geometry, this must have been the survival mechanism. Cell manipulation naturally led to the ability to specialize different parts of the body for different activities. Then, some abnormal terrene of cells developed the ability to create new globes and affix them to the original terrene. These activities resulted in the formation of Porifera, the sponges. The sponges have no particular shape; this may have been attributed to the manipulation of the cell-to-cell interactions (Palaeos.com 2002).
Ernst Haeckel (1834-1919) in his animal research was the first person to recognize the astonishing distinction between early animal development and the feasible course of early animal evolution. He called christened his idea the “Gastraea Hypothesis” and therefore the origin of Haeckel’s “Law“ that stipulates the “ontogeny recapitulates phylogeny.” Haeckel’s Law It is not actually a “law” but a recurring pattern. It explains animal evolution without giving scientific evidence as to why some things happen (Palaeos.com 2002).
Urbilateria classifications include all Metazoa which has bilateral symmetry. A bilateral symmetric animal means that the animal body is comprised of a front and rear, the left and the right body parts during their development. In the course of development, many bilateria display cephalization characteristics that draw them to concentrate on developing sensory structures such as the mouth, nerve ganglia, and the anterior end. Bilateria are also triploblastic and they develop three germ layers during embryonic development. The three germ cells include Ectoderm, Endoderm, and Mesoderm. Ectoderm germ cell covers the surface of the embryo and on maturity, it covers the animal’s outer cover and forms the central nervous system in some phyla. Endoderm forms the central part of the germ layer which lines the archenteron, the digestive system, and another pocket-like projections that forms the liver and lungs of vertebrates. The mesoderm, the central part, is located between the ectoderm and endoderm. Mesoderm forms the muscles and organs systems situated between the digestive tract and outer layer of the animal. It serves as a circulatory system and development of the skeletal system (Palaeos.com 2002)
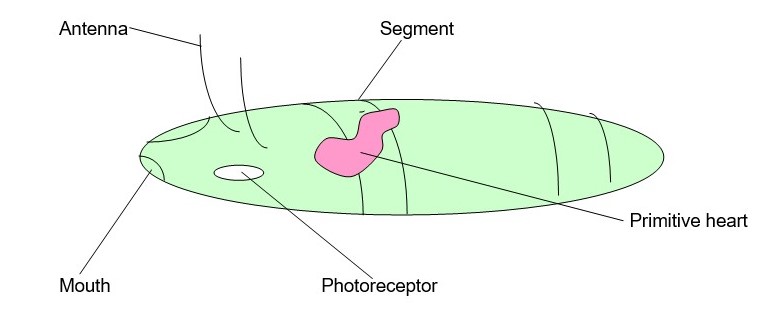
Past studies on Urbilateria development believed that the last ancestors of these animals were segmented that reflects the recent evidence from comparative molecular embryology development. The genetic evidence reveals that the morphological and developmental of Urbilateria segmentation emerged from the molecular phylogeny of metazoans. Urbilateria segmentation characteristics are colossally complicated as no clear definition has clarified the segmentation terminology and as well as its taxa group. Balavoine and about (p.2) argue that Urbilateria ancestral segmentation might have undergone many complex etymologies in each different phylum making its cladistic mapping problematic.
In the general definition of Urbilateria segmentation, some remarkably similar patterns are found across the bilaterian tree that explains the way segments are formed. This explains that segmentations are formed by the posterior addition of mesodermal segments or somites. It’s therefore assumed that the strong similarities portrayed in mesodermal patterns are ancestral, as the features display bilaterians from a metameric ancestral resemblance. Several types of evolutionary mechanisms attributed to body segmentations such as specialization, tagmosis, and progenesis would explain the rapid emergence of body plans during Cambrian. Since the researcher has come up with a clear definition of Urbilateria segmentation, Balavoine and about (2) propose a hypothesis test that will compare genes involved in mesodermal segmentation.
The recent research advances on animal segmentation concerning the phylogeny of metazoans and the identification of their ancestral group of conserved patterning genes have generated a stimulating debate about the morphology of the last common ancestor of the Urbilateria. It also studies how the astonishing dissimilarities of the bilaterian body plans came into existence. In the past years, some molecular embryology data on amphioxus have reawakened the hypothesis that the ancestor of coelomates was a segmented animal. On the concept of the molecular phylogeny of metazoans, the earlier studies have shown Urbilateria to be a complex organism, coelomate, and consist of segmented body parts. Balavoine & Adoutte (p.2) therefore tries to develop a more detailed study on the morphology, development, and phylogenetic arguments that countenance the segmentation of bilaterian ancestor. All these genetic arguments do not clearly define the developmental genetic evidence for homology illustration. They, therefore, attest to the ambition of studying the similarities between the morphological and developmental patterns in distant phyla. Their studies were efficient enough to be considered in the hypothesis of a metameric Urbilateria and to be used in future research in the study of possible axes of comparative embryology.
As we continue with Urbilateria characteristics, we discover the shared roles of Pax6 and Six homologs in the eye development that takes us back to the phenomena of Bilateria common ancestries that were characterized by the simple form of eyes. In Urbilateria eye development, the homology of bilaterian cerebral eyes at the level of eye anatomy consists of cell types and phototransduction molecules. One of the bilaterian eyes is known as the larval pigment-cup eyes and they are located to the left and right of the apical organ which is the primary ciliary larvae of Protostomia and Deuterostomia bilaterians. The second eye classification consists of a single pigment cell and a single photoreceptor cell in inverse orientation. The third classification is more elaborate and consists of cerebral pigment-cup eyes with an reverse arrangement of photoreceptor cells mostly found in adult Protostomia. Both inverse larval and reverse adult eyes consist of rhabdomeric photoreceptors but the distinguishing factor in their eyes is the difference in the chordate cerebral eyes which shows ciliary photoreceptors. The difference is significant due to the molecular level for phototransduction rhabdomeric and ciliary photoreceptor cells. phototransduction rhabdomeric employs divergent rhodopsins while ciliary photoreceptor cells employ non-orthologous G-proteins. Distinguishing factor also occurs where phototransduction rhabdomeric uses rhodopsin kinases and ciliary photoreceptor cells use arrestins. This comparison helps in the support of homology of cerebral eyes in Protostomia and does not back the homology of chordate and non-chordate cerebral eyes that uses photoreceptor cells with non-orthologous phototransduction cascades (Arendt & Wittbrodt 2008).
In conclusion, Haeckel law does not provide any concrete evidence on the development of Urbilateria segmentation; it should therefore not be used as a study in animal evolution. The research provides a conflicting definition of what Urbilateria is, we therefore conclude Urbilateria to be the ancestor of all bilateria and its physical characteristics would be a worm-like animal with a mouth, a primitive gut and has sensory structures. Internally, its known to have body wall muscles and is tripolastic in nature.
References
- Balavoine Guillaume $ Adoutte Andre. “Integrative and Comparative Biology.” The Segmented Urbilateria. A Testable Scenario 43 (2003): 1-11.
- Palaeos.com. 2002. Metazoa: the Animals.
- Arendt, D & Wittbrodt, J.” Reconstructing the eyes of Urbilateria.)”. PubMed Central (PMC3 – NLM DTD (2008)