General Description of Homo Floresiensis
Homo Floresiensis is a primitive hominine species that has become an object of active scientific debates. H. Floresiensis was discovered in 2003 in Lang Bua (Indonesia) in the Late Pleistocene sediments (Sutikna et al., 2016). For a long time, the scientists have been discussing the controversial questions concerning this species. First of all, it was unclear whether it had been an extinct Homo species, or a modern human that has migrated and pathologically changed. In this paper, the questions of distinctive features and discovery of H. Floresiensis will be analyzed.
Discovery of Homo Floresiensis
The bones of H. Floresiensis were first discovered during the excavations that took place from 2001 to 2004 in Lang Bua cave, Indonesia, on the Flores island. The skeletal remains were found in the Late Pleistocene sediments at the depth from 4 to 7 m in different excavated areas (Sutikna et al., 2016). The discovery was made in 2003 by a team of Indonesian and Australian archaeologists.
Culture of H. Floresiensis
H. Floresiensis has used a range of primitive stone tools. The tools are similar to Oldowan ones and are represented by blades, perforators, and points (Baab, 2012). The instruments could be used for killing Stegodons, extinct elephants whose fossil remains were also found in the same cave with H. Floresiensis. Similar artefacts that were found in the Soa Basin cave, 50 km south, are attributed to H. Erectus (Baab, 2012). Thus, H. Floresiensis could use instruments despite its small brain volume.
Ancestry of Homo Floresiensis
The ancestry of H. floresiensis is a matter of controversy. According to one hypothesis, it has evolved from Asian H. Erectus (van der Bergh et al., 2016). In this case, it can represent an example of evolution among hominines put in an isolated environment. Sutikna et al. (2016) assume that H. Floresiensis could have been isolated during the Wisconsin glaciation. Thus, it can serve as a prove for evolution theory, as the isolated species have changed greatly.
According to the alternative point of view, this taxon has derived from Homo habilis or even from Australopithecus. This can mean that the hominines have “dispersed from Africa to Eastern Asia by two million ago” (van der Bergh et al., 2016, para. 7). However, the closest species recognized as H. floresiensis ancestor and discovered in Mata Menge is derived compared to H. Habilis and Australopithecus (van der Bergh et al., 2016). It can support the hypothesis that H. Floresiensis is still a descendent of Asian Homo habilis, although extremely dwarfed.
However, there are no skeletal remains that would prove the idea that there is a deeper ancestor of H. Floresiensis than H. Erectus. The only before H. erectus skeleton remains were found in Africa. Besides, there are not so many records of the hominines that preceded H. Erectus (Baab et al., 2012). This fact makes it difficult to compare the pre-Erectus fossils to H. Floresiensis. Thus, the ancestry of the species is unclear, as well as the time of its derivation. The probable representation of the hominine family tree is presented in Figure 2.
Distinctive Features of Homo Floresiensis
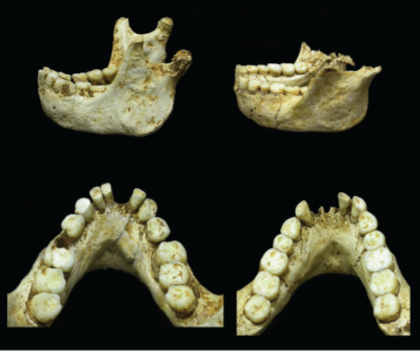
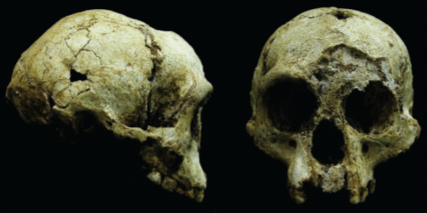
H. Floresiensis has a number of distinctive features that have become an object of discussion among the scientists. It has a small volume of brain (about 417 cm3), short stature (from 1.00 to 1.09 m), and features that “appear phylogenetically primitive” compared to modern humans (Baab et al., 2016, p. 1). Due to the small body size, this taxon is also known as “Homo Hobbitus” or just the “Hobbit”. These features differ the species from other hominines, although there are some questions to be discussed. For example, H. Floresiensis has small teeth, like other hominins, which allows assuming that this is a distinct hominin species tracing back to more than 700,000 years (Gomez-Robles, 2016). According to Van der Bergh, teeth of such size are typical for Homo Sapiens (Van der Bergh, 2016). The cranes and the jaws of H. Floresiensis are presented in Figure 1.
The Down Syndrome Hypothesis
However, other researchers argue that H. Floresiensis represents modern human species. In particular, it is claimed that H. Floresiensis is a modern Austro-Melanesian hominine with the signs of Down syndrome (Baab et al., 2016). Baab et al. have carried out a number of researches, including endocranial measurements, analysis of MRI data, analysis of neurocranial and mandibular symphysis shape, relative premolar length, etc. The results revealed that the cranial vault of Homo Floresiensis sharply contrasts with that of modern humans, including the patients with Down syndrome. The cranial vault of H. Floresiensis is more similar to H. Erectus, as its “cranial base is not flat” (Baab et al., 2016). Besides, the angle of its cranial base “overlaps the euploid but not the Down syndrome range of values” (Baab et al., 2016, para. 12). In the whole, the cranial morphology of H. Floresiensis is more common in archaic Homo than in the modern one.
Evolution Theory
The small stature of H. Floresiensis can support the evolution theory. If the hypothesis about its derivation from H. Erectus is right, then H. Erectus has adapted to the isolation on the island by dwarfing. Another species living on islands also tend to have small stature. This phenomenon has been also recorded in deer, mammoths, and primates (Baab, 2012). It seems that small stature helped the species to adapt to the limited volume of resources on an island.
Conclusion
To conclude, it can be said that the ancestry of H. Floresiensis is unclear. There have been several theories concerning their derivation from other hominines. Some researchers believed it to be a modern human with the signs of Down syndrome. However, the prevailing theory suggests that H. Floresiensis has acquired its distinctive features as a result of isolation on the island, which supports evolution theory.
References
Baab K. (2012) ‘Homo floresiensis: making sense of the small-bodied hominin fossils from Flores‘. Nature Education Knowledge, 3(9).
Baab, K. L. et al. (2016) ‘A Critical evaluation of the Down Syndrome diagnosis for LB1, type specimen of Homo floresiensis‘, PLoS One, 11(6), pp. 1-32.
Gomez-Robles, A. (2016) ‘The dawn of Homo floresiensis‘, Nature, 534(7606), p. 188.
Sutikna, T. et al. (2016) ‘Revised stratigraphy and chronology for Homo floresiensis at Liang Bua in Indonesia’. Faculty of Science, Medicine and Health – Papers. Web.
van den Bergh, G. D. et al. (2016) ‘Homo floresiensis-like fossils from the early Middle Pleistocene of Flores’, Nature, 534(7606), p. 245. Web.