Introduction
Chadha et al (1982) claims human Interferon alpha is protein derivative that is biosynthesized and secreted by lymphocytes under pathogenic stimulus. Allen and Diaz (1996, p.182-3) argues that the main stimulus that predispose secretion of human alpha are viruses, bacteria, parasites or tumor cells. Spiegel (1989, p.76-77) claims human interferon alpha provides foundation for communication between leucocytes hence facilitates in immunological functions. The classification of interferon alpha has been documented by Allen and Diaz (1996, pp.181-184) to belong to group of specialized compounds known as glycoprotein that are termed as cytokines. Ozato et al (2007) suggested that Glycoprotein consists of two macromolecules made of carbohydrates and proteins. Allen and Diaz (1996, p.181) has demonstrated that nomenclature of interferon alpha is based on its functionality through “interference” with replication or biological processes of pathogens. Other biological roles of interferon alpha include activation of immune system (Ozato et al, 2007), stabilization of healthy cells such that they can resist pathogenic infections.
Interferon cellular production
Berg et al (1982, pp.23-6) claims that as a pathogenically infected cell undergoes lysis or death from cytolytic pathogens influence, it releases pathogenic particles that catalyze progression and propagation of infection. However, Chadha and Sulkowski (1985) argument is based on assumption that the affected cell signals other healthy cells by secreting or releasing interferon alpha as documented by Klaus et al (1997). The healthy neighboring cells through a negative feedback process as claimed by Isaacs and Lindenmann (1957, 1987) secrete a counter-enzyme termed as Protein-Kinase-R (PKR) whose primary biological role is to phosphorylate a protein termed as eIF-2 (Chadha et al (1982).
According to Chadha and Sulkowski (1985, pp.45-51), eIF-2 is responsible for eukaryotic translation initiation that results into formation of inactive complex via eIF-2B which decreases protein biosynthesis hence interfering with biochemical processes that can support bioactivity of the pathogen. Allen and Diaz (1996, p.182) claims another enzyme termed as RNAse-L initiations infected cell destruction which can either occur through lysis or force the cell to commit suicide through apoptosis which terminates protein biosynthesis of the pathogen and the pathogen-host cell. Interferon alpha has been documented in literature (Chadha et al, 1982; Berg et al, 1982; and Isaac and Lindenmann, 1987) to initiate production of specific proteins known as Interferon-stimulating Genes (ISGs) that have pathogen receptors and destroy the pathogen. Interferon alpha according to Spiegel (1989, pp.76-7) increase P53 activity which reduce spread of the pathogen and induce cell apoptosis
Aims of the essay
The essay reports on purification and isolation of human interferon alpha protein. The essay seeks to report on the first method of purification and isolation of interferon alpha that was used, provide biochemistry of separation steps that were used and form a flow chart based on the initial interferon alpha purification and isolation. The essay reports on successful methods and unsuccessful methods that were adopted and rationale for the adoption of the methods. The essay reports on the processes that were used to precipitate the interferon alpha (salting out) and methods of chromatography that were applied.
The discovery of the human interferon alpha protein
Allen and Diaz (1996) and Berg et al (1982) claim that two hypotheses have been put forward to demonstrate mechanism through which human interferon alpha was discovered.
First proposed method towards discovery of interferon alpha protein
In literature (Isaac & Lindenmann, 1987, pp.429-438) claim interferon alpha was discovered in 1957 in Chickens by Alick Isaac and Jean Lindenmann who were attached to the National Institute for Medical Research in London. The two virologists observed that there was interference effect that arose from heat-inactivated influenza virus (Allen & Diaz, 1996) that occurred on live influenza virus that was cultured in a chicken egg Chorioallantioc membrane (Berg et al, 1982). Isaacs and Lindenmann (1987) claims the results for discovery of the interferon alpha were published in 1957. Due to interference effect Isaacs and Lindenmann (1987) named the protein “interferon” based on its property of interference effect. Allen and Diaz (1996) have identified the interferon alpha as Type I Interferon. Follow up studies in National Institute for Medical Research (1978), determined that human being produced interferon alpha through the same biochemical mechanism. The first isolated and purified human interferon alpha was carried out in 1981. Increased research on biochemistry of Type I interferon (Allen & Diaz, 1996) between 1978 and 1981 resulted into determination of protocol for purification and isolation of Type I interferon alpha and Type I interferon beta (Klaus et al, 1997).
Second proposed method towards discovery of interferon alpha protein
The second proposed discovery of interferon alpha has been reported to have occurred earlier before Isaacs and Lindenmann (1987) discovered interferon alpha in 1957 (Isaacs & Lindenmann, 1957). The discovered was pioneered by Yasu-Ichi Nagano and Yasuhiko Kojima, Japanese virologists in 1954 (Nagano & Kojimo, 1954). The Japanese virologists were residents at the institute For Infectious Diseases at university of Tokyo. The virologists were working on vaccine for small pox when they observed that there was inhibition of viral growth in an area of a rabbit skin that had been inoculated with Ultra-Violet inactivated virus (Isaacs & Lindenmann, 1957).
In conclusion (Nagano & Kojimo, 1954) claimed that there was a virus inhibiting factor in the rabbit tissue. Nagano and Kojimo (1958) conducted series of research towards isolation, purification and characterization of the interferon alpha. The findings of Nagano and Kojima were published in a French journal named “Journal de la Société de Biologie” in 1954 (Nagano & Kojimo, 1954). The Japanese virologists carried out further studies and determined that the viral activity of the virus-inhibiting factor had a lifespan of 1-4 days and was independent from antibody production. Nagano and Kojima findings (Nagano & Kojima, 1954; 1958) determined that the virus-inhibiting factor had no relationship with antibody production were published in 1958 (Nagano & Kojimo, 1958) a year after Isaacs and Lindenmann (1957; 1987) published their findings and named the “virus-inhibiting factor” as Interferon alpha.
How interferon alpha was purified initially purified
The interferon alpha was isolated and purified through centrifuging and reverse phase high performance liquid chromatography (RP-HPLC) (Isaacs & Lindenmann, 1957). The procedure involved loading the crude interferon alpha into a glass absorbent chromatography, followed by eluting interferon alpha from the glass absorbent by using standardized Hydrophobic electrolyte that had a PH range of 7-8.5. This was followed by loading the interferon alpha eluate through a molecular sieving chromatography which resulted into a resolution that had a range of 10k to 100K IU (Nagano & Kojimo, 1954; 1958). This was repeated but with a different hydrophobic electrolyte until viruses was eliminated. Finally the eluate was entered into a Zinc II ions chelate resin which resulted into collection of non-adsorbed eluate that had purified interferon alpha.
Discovery route for interferon alpha
Detailed protocol for purification as a flow chart.
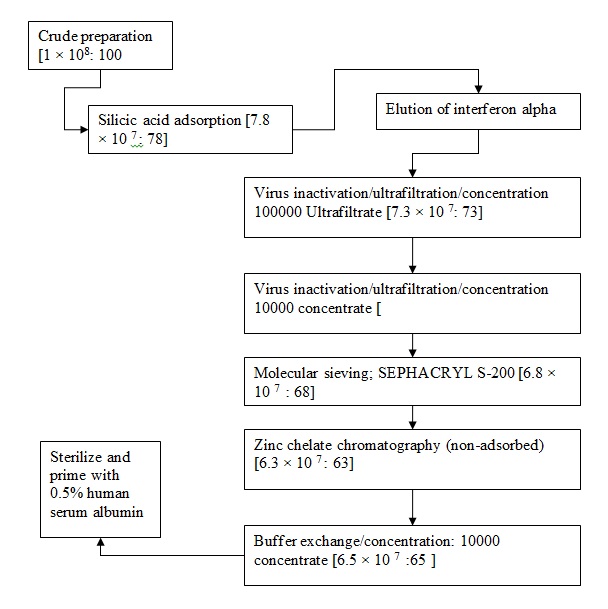
Unsuccessful methods for purification
The procedure for purification of the interferon alpha of successful purification
A batch that contained crude interferon alpha that was determined to have 10000 IU in terms of MDBK cells was adopted for a batch-to-batch absorption via use of 15ml of silicic acid that had been equilibrated by using 20.00mM phosphate buffer whose PH had been standardized to 7.4 (Isaacs & Lindenmann, 1957). The adsorption was allowed to progress for 90 minutes at a temperature of 4oC as the mixture was stirred. The gel that resulted was put into a glass column that measured 0.9cm by 25cm and immediately washed by using 100ml of 20.00mM phosphate buffer whose PH had been standardized at 7.4 (Nagano & Kojimo, 1954; 1958). interferon activity was recovered from the silicic acid by using 100.00 mM TRIS-HCL whose PH had been standardized to 8.0 to achieve 205 interferon alpha activity followed by 100.00 mM TRIS-HCL that contained 0.5 Molar TMAC, 0.5 Molar Sodium Chloride and 10% propylene glycol whose PH had been standardized to 8.0 to achieve 70% interferon activity (Isaacs & Lindenmann, 1957). A 10% remaining activity was however detected in the non-adsorbed fraction of the silicic acid which was subject to presence of sendai virus residuals (Hauschild et al, 2008).
Successful methods for purification
The procedure for purification of the interferon alpha of successful purification.
Step one: silicic acid adsorption
A sample that has been determined to contain interferon alpha is taken in its crude state and cooled at 4oC before purification procedures are started (Isaacs & Lindenmann, 1957; Nagano & Kojimo, 1954). The step involves a batch-to-batch operation as claimed by Isaacs and Lindenmann (1957). After interferon alpha is cooled, it is mixed with silic gel (Liu, 2005). The preferred silica gel is silicic acid. Alternatively, crude cooled interferon alpha is introduced into a controlled pore glass (CPG) as claimed by Isaacs and Lindenmann (1957). The CPG should have been activated with 20.00 mM sodium phosphate buffer solution which should have a PH of 7.4 (Nagano & Kojimo, 1954). The ratio of the silicic acid to the culture solution of interferon alpha should be within the range of 1:10 to 1:50. In literature, a ratio of 1:30 has been proposed to be adequate as documented by Isaacs and Lindenmann (1957) and Nagano and Kojimo (1954; 1958). The mixture should be stirred at a low speed for about 60 minutes to 90 minutes. The adsorption temperature should be thermoregulated at 4oC (Isaacs & Lindenmann, 1957). The gel should be transferred into a glass column after 60-90 minutes as claimed by Isaacs and Lindenmann (1957). In the glass column, the gel should be washed by using 20.00mM phosphate buffer solution whose PH should be standardized to 7.4 (Sen, 2001). The washing with the buffer should be continued until adsorption at 280.00nM attains a background level.
Elution of the interferon alpha
The elution of interferon alpha should be conducted via use of a buffer that contains 100.00 mM TRIS-HCL, 0.5Molar solution of Sodium chloride and 0.5 Molar solution of Tetramethylammonium Chloride (TMAC) and 10% concentration propylene glycol with PH standardized at 8.0 (Isaacs & Lindenmann, 1957). Optimization of the elution buffer, as claimed by Isaacs and Lindenmann (1957) ensures dissociation conditions are provided which decrease opportunities for formation of complexes hence realization of higher yield of interferon alpha inter-species (Nagano & Kojimo, 1958). The elution of interferon alpha results into a five times concentration and removal of utmost 93% of proteins as documented by Isaacs and Lindenmann (1957)
Inactivation of the virus
This step was meant to realize virus, virus inactivation and concentration (SEn, 2001).
Virus inactivation was carried out through ultrafiltration and concentration determined (Nagano & Kojima, 1954). This was carried by using eluate that contained interferon alpha activity mixed with non-ionic detergent TRITON X-100 to achieve a concentration of 0.1% TRITON X-100. In absence of TRITON X-100, other non-ionic detergents can be used for instance TWEEN-20 and NONIDET P-40 as claimed by Isaacs and Lindenmann (1957). Gentle stirring should be carried out continuously for thirty minutes at a temperature of 4oC which results into destruction any present viruses (Nagano & Kojima, 1958). Post incubation, the product should be transferred onto a Millipore cassette which could preclude flow of proteins or compounds that had molecular weight that was in excess of 100000 (Isaacs & Lindenmann, 1957).
The ultrafiltrate that is achieved contains proteins that have molecular weight that is less than 100000 which further undergoes repeated stirring through ultrafiltration system with the membrane whose pore sizes are precludes to allow flow of materials that have molecular weight that is less than 10000 (Isaacs & Lindenmann, 1957). This is step is repeated until the right concentration is achieved. The aim of repeated ultrafiltration is to ensure all viruses are removed in case there are some viruses that might have been resistant to non-ionic detergents (Nagano & Kojimo, 1954; 1958 and Isaacs & Lindenmann, 1957). The final concentrate consists of proteins that have a molecular weight of 10000 and 100000 IU which proceed to the next phase that involves use of molecular sieving.
Molecular Sieving
The concentrated material that contained proteins whose molecular weight ranged between 10000 and 100000 was loaded into a 12cm by 180cm column that was impregnated with SEPHACRYL S-200 (Isaacs & Lindenmann, 1957). In the second trial, sieving beads that had a resolution that ranged from 10000 to 100000 molecular weights were used. The molecular sieving was conducted at a temperature of 4oC in presence of 10mM phosphate buffer, 0.5Molar solution of sodium chloride and 10% glycol at a standard PH of 7.4 (Hauschild et al, 2008; Isaacs & Lindenmann, 1957). The proteins were eluted depending on their molecular weight. Different fractions were achieved that were transferred into a Zinc II ions chelate agarose chromatography (Isaacs & Lindenmann, 1957).
Zinc Chelate Chromatography
The product of molecular sieving column was applied on a column that was impregnated with a chelating matrix (Isaacs & Lindenmann, 1957). The Zinc chelating chromatography used a Zinc II ions Chelate agarose. After loading of the combined product from the molecular sieving stage Isaacs and Lindenmann (1957) claim, the column containing the chelating matrix was washed with 10 mM phosphate buffer that further contained 0.5Molar solution of sodium chloride whose PH had been standardized to 7.4. The procedure was carried out at a temperature of 4oC (Navratil et al, 2010).
Buffer Exchange/Concentration
Alick and Lindenmann (1957) claims non-adsorbed material is concentrated and buffer is exchanges to 20mM phosphate buffer at PH of 7.4. This is carried out by use of ultra-filtration subject to use of a membrane that is impermeable to protein that has a molecular weight of above 10000 (Fensterl & Sen, 2009). The process of ultra filtration is carried out until compete buffer exchange is achieved that is determined by concentration of interferon of about 100,000 to 100,000 IU (Isaacs & Lindenmann, 1957). This is followed by addition of human serum albumin which is used as a stabilizing agent for the interferon alpha. The human serum albumin concentration should be within the range of 1miligram to 10 milligrams per milliliter (Nagano & Kojima, 1954; 1958). The product of stabilization should be filtered through a 0.22 uM filter (Liu, 2005). It should be noted that the filters should be primed with 0.5% human serum albumin before sterilization which ensures the interferon alpha does not lose its activity (Isaacs & Lindenmann, 1957; Liu, 2005)).
Diseases that are managed by interferon alpha
Some of the diseases that are treated by using interferon alpha include hairy cell leukemia, genital warts, Aids related Kaposi Sarcoma, Non-A hepatitis, Non-B hepatitis and Hepatitis B (Fensterl & Sen, 2009).
Diseases caused by deficiencies in enzymes
Navratil et al (2010) claims deficiencies or disorders in enzymes result into diseases that are characterized by loss of lysosomal storage for instance Sly Syndrome that is a Type MPS VI and characterized by loss of Lysosomal storage. Lysosomal storage diseases have been documented in literature to be associated with functionality of glycosaminoglycans (Fensterl & Sen, 2009). The disorders are characterized by accumulation of glucuronic acid containing glycosaminoglycans (dermatan, heparin and Chondroitin 4-and 6-sulfates. Other types of MPS include MPS I H or Hurler syndrome, MPS I S or Scheie syndrome, MPS I H-S or Hurler-Scheie syndrome, MPS II or Hunter syndrome, MPS III or Sanfilippo syndrome, MPS IV or Morquio syndrome, MPS VI or Maroteaux-Lamy syndrome and MPS VII or Sly syndrome, MPS III or Sanfilipo Syndrome has other subtypes for instance Sanfilippo B that is caused by deficient enzyme alpha-N-acetylglucosaminidase; Sanfilippo C that is caused by altered enzyme acetyl-CoAlpha-glucosaminide acetyltransferase and Sanfilippo D that is caused by deficient enzyme N-acetylglucosamine 6-sulfatase.
References
Allen G, Diaz MO. (1996) “Nomenclature of the human interferon proteins”. J Interferon Cytokine Res; 16: 181–184.
Berg et al., (1982) “Purification and Characterization of the HuIFN-a Species,” Texas Reports on Biology and Medicine, vol. 41.
Chadha and Sulkowski, (1985) “Production and Purification of Natural Human Leukocyte Interferons,” The Interferon System; A Current Review (New York), Kruzel et al., Oct.
Chadha et al. (1982) “Adsorption of Human Alpha (Leukocyte) Interferon on Glass: Contributions of Electrostatic and Hydrophobic Forces,” Journal of Interferon Research, vol. 2, No. 2
Fensterl, V; Sen GC (2009). “Interferons and viral infections”. Biofactors 35 (1): 14–20.
Hauschild, A.; Gogas, H.; Tarhini, A.; Middleton, M.; Testori, A.; Dréno, B.; Kirkwood, J. (Mar 2008). “Practical guidelines for the management of interferon-alpha-2b side effects in patients receiving adjuvant treatment for melanoma: expert opinion”. Cancer 112 (5): 982–994.
Isaacs A, Lindenmann J (September 1957). “Virus interference. I. The interferon”. Proc. R. Soc. Lond., B, Biol. Sci. 147 (927): 258–67.
Isaacs A, Lindenmann J. (1987) “Virus interference. I. The interferon”. J Interferon Res; 7: 429–438.
Klaus W, Gsell B, Labhardt AM, Wipf B, Senn H. (1997) “The three-dimensional high resolution structure of human interferon alpha-2a determined by heteronuclear NMR spectroscopy in solution”. J Mol Biol; 274: 661–675.
Liu YJ (2005). “IPC: professional type 1 interferon-producing cells and plasmacytoid dendritic cell precursors”. Annu Rev Immunol 23: 275–306.
Nagano Y, Kojima Y (1958). “Inhibition de l’infection vaccinale par un facteur liquide dans le tissu infecté par le virus homologue” (in French). C. R. Seances Soc. Biol. Fil. 152 (11): 1627–9.
Nagano Y, Kojima Y (October 1954). “Pouvoir immunisant du virus vaccinal inactivé par des rayons ultraviolets” (in French). C. R. Seances Soc. Biol. Fil. 148 (19-20): 1700–2.
Navratil V, de Chassey B, et al. (2010-11-05). “Systems-level comparison of protein-protein interactions between viruses and the human type I interferon system network.” Journal of Proteome Research 9 (7): 3527–36.
Ozato K, Uno K, Iwakura Y (May 2007). “Another road to interferon: Yasuichi Nagano’s journey”. J. Interferon Cytokine Res. 27 (5): 349–52.
Sen GC (2001). “Viruses and interferons”. Annu. Rev. Microbiol. 55: 255–81.
Spiegel RJ. (1989) “Alpha interferons: a clinical overview”. Urology; 34: 75–79.