Introduction
Symbiosis between marine Cnidarians (coral reefs; Order Scleractinia) and unicellular photosynthetic dinoflagellates (algae; genus Symbiodinium) is a common feature of marine biota. While Cnidarians feed on zooplankton and supply the dinoflagellates with assimilable nitrogen, phosphorus and carbon, in turn the dinoflagellates, commonly known as zooxanthellae, derive photosynthates notably glycerol, glucose and some amino acids for the animal partner (Clayton & Laskerh, 1984). Cnidarians bleaching is a deterrent phenomenon that severely disturbs the coral reef ecosystems, caused by various stressors like heavy metal and other pollutants, high irradiance, ultraviolet exposure, low salinity and high temperature. A temperature barely 1-2oC above the summer tropical water surface temperatures is sufficient to induce Cnidarian bleaching. According to Ralph, Gademann & Larkum (2001) the optimal surface water temperature of coral reef stays around 28.5oC, and incidences when temperature was risen to ca. 32oC about 80% of reef were destructed by bleaching. Contrary to this most of the marine organisms are rarely affected to temperature rises even up to 40-45oC. Temperature-induced Cnidarian bleaching is either a collapse of symbiotic interaction or is a loss of photosynthetic pigments- chlorophyll and carotenoids of the dinoflagellates. Evidence suggest that several host factors viz. protein hyperphosphorylation, calcium deposition, reactive oxygen (nitrous oxide) production etc. are accountable for this phenomenon (Sawyer & Muscatine, 2001; Weis, 2008).
We tested a hypothesis that more than actual pigment loss it is the detachment and population reduction of zooxanthellae that contributes to Cnidarian bleaching. Since zooxanthellae are also associate with sea anemone Aiptasia pulchella, we selected it as test model and monitored change in zooxanthellae counts and chlorophyll content as a function of time in two higher than optimal temperature regimes.
Material and Methods
We have collected Aiptasia pulchella individuals from their natural habitat and set up growth conditions in two tanks (capacity?). We placed about 10 animals in distilled water, to which Instant ocean salt mix equivalent to 35 g L-1 was added (35 ppt salinity). The temperature was set at 28oC and 32oC. One additional tank kept at 25oC was used as control. The water was periodically circulated for aeration and as feed small particle fish food from local market was given two/ three times a week (check?). We have sampled three animals at zero time from the control tank and data was used for all the other sets. Further, we have sampled 5 animals from the treated sets (higher temperatures) on first and third week from the respective tanks. The polyps were weight (in mg) after being dried with paper towel. We then homogenized in 5 ml of seawater using a hand homogenizer. Large polyps we cut in two halves before homogenizing. We then separated zooxanthellae by vortexing the samples for 1 min and divided the suspension in two parts. Using 1 ml sample we proceeded for chlorophyll estimation by vortexing with 9 ml of acetone. After settling the coarse precipitates the samples were centrifuged at 1000×g for 5 min and the supernatant was decant off. The absorbance was recorded at 665, 647, and 630 nm against acetone blank. We calculated chlorophyll concentration using the formula (A);
Chlorophyll (µg) = [(11.85A665) – (1.54A647) – (0.08A630)] × [5 (ml homogenate)/0.5]
Thereafter, we applied a drop of the polyp homogenate to the hemocytometer chamber for counting under high power (450 X). Each counting chamber was square of area of 3 mm enclosing 3 mm × 3 mm = 9 sq. mm area, which was divided in to nine squares each having 1 sq. mm area. The central square was divided in 25 small squares so each of these small squares had 1/25 sq. mm area, which was further divided in 16 small squares each of which now had 1/25 × 1/16 = 1/400 sq. mm area. We counted zooxanthellae in 5 of the 25 small squares. This means, counting of the cells was done in 5 × 16 = 80 small squares having a volume of 1/400 × 1/10 = 1/40003 mm of the sample per sq. Hence, no. of cells (B) = [(Total no. of cells counted × dilution × 4000)/ no. of small squares] × 5 (ml homogenate). We also calculated chlorophyll/ zooxanthellae ratio as (A) ÷ (B).
Results
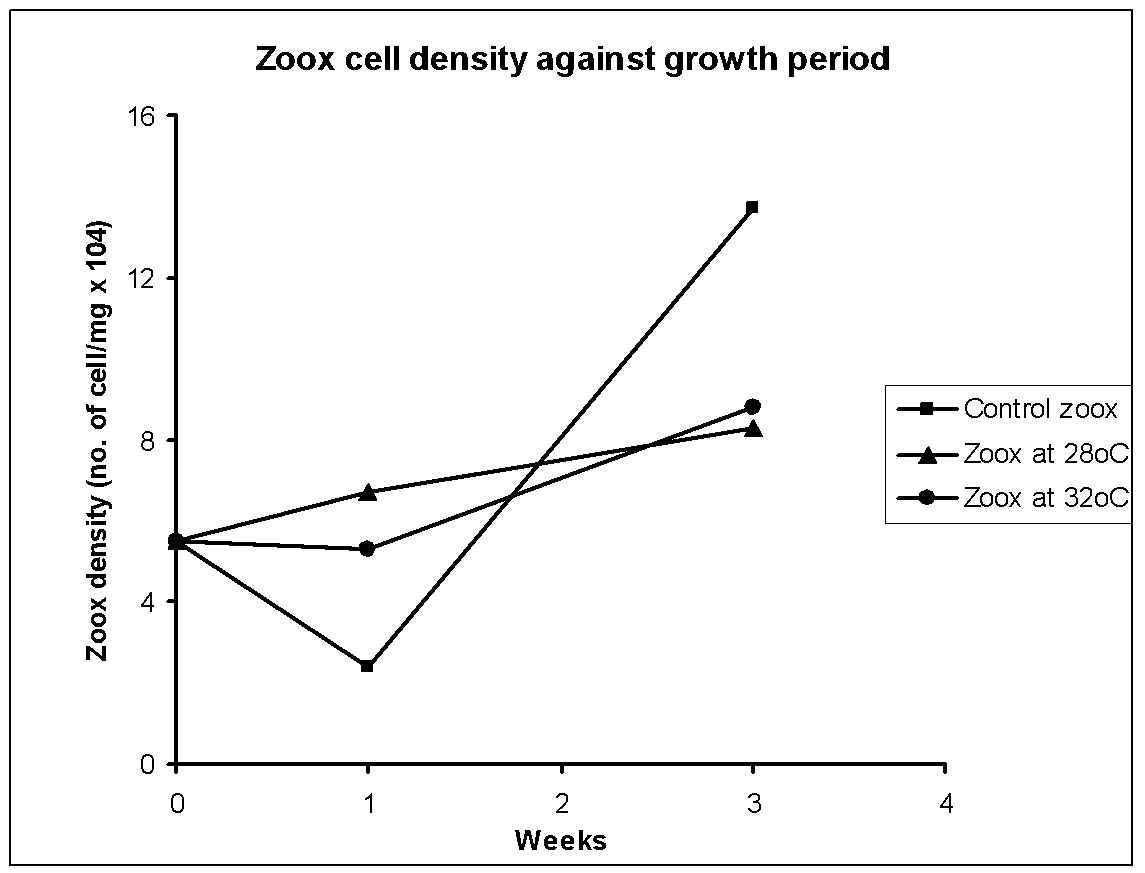
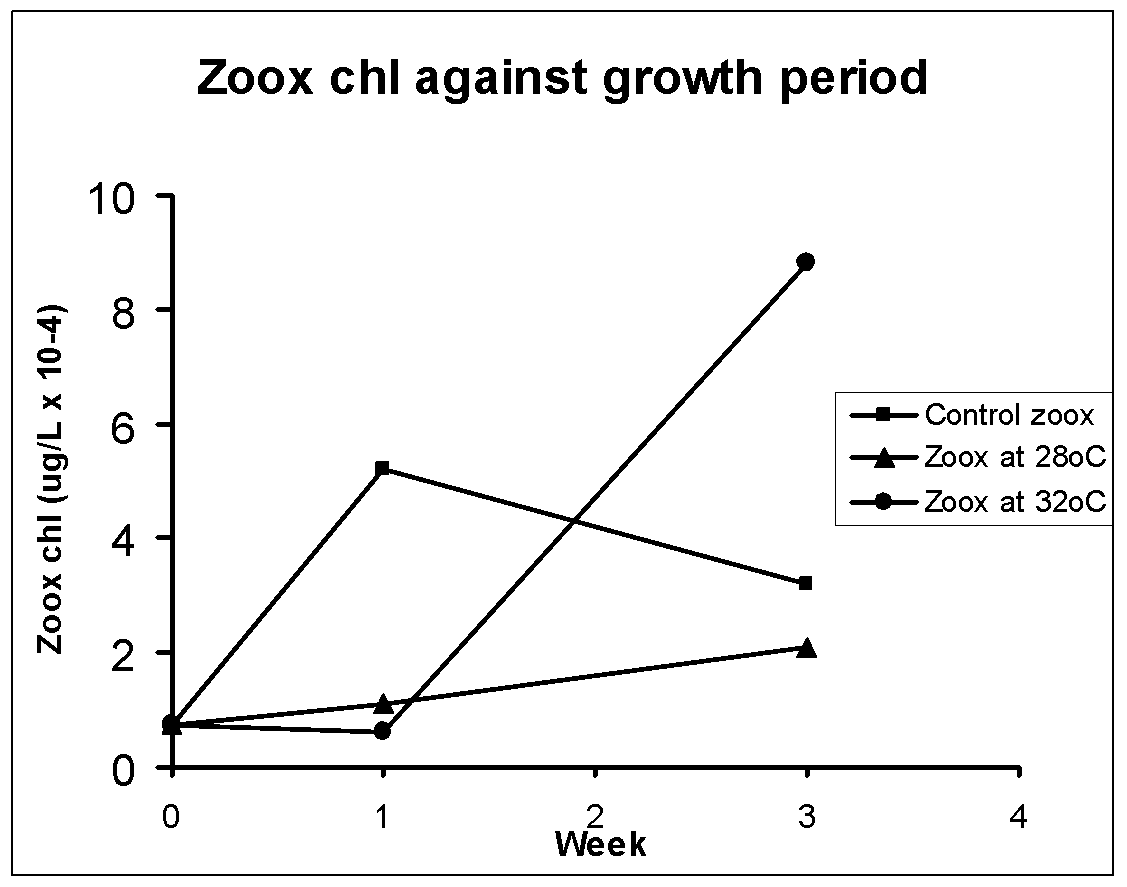
The average values of zooxanthellae cell number per mg animal and chlorophyll per liter in the control (n=3) and two high temperature sets (28, 32oC; n=7-10) of homogenized sea anemone is shown respectively in Fig. 1 and 2. In control the initial number declined and thereafter increased, but chlorophyll was high during first week and then it kept decreasing. The zooxanthellae counts and chlorophyll in animals kept at 28oC steadily increased though this increase was much lower than the controls in prolong incubation. Somewhat same increase in zooxanthellae number was recorded even at 32oC but here chlorophyll content did not change until 1 week and thereafter it shot up to a level even higher than control. Hence overall growth related changes in cell number and chlorophyll content did not match each other which indicate that the parameters responded differently to growth period and to the tested temperatures.
Discussion
There was a completely opposite trend in number and chlorophyll content in the controls during the given growth period. This suggests that zooxanthellae was still in the acclimatization phase when lower numbers of cells had excess chlorophyll in the beginning and later when cells were acclimatized the cell density sharply increased though there was not much increase in chlorophyll content. The hypothesis that zooxanthellae dislodged at high temperatures can be justified if during growth at 28/32oC both zooxanthellae count and chlorophyll content would have concomitantly decreased compared to the control. If we consider the week 3 chl data of 32oC as experimental artifact, then we indeed found lower chlorophyll and cell number in heat treated samples than the controls during the growth phase. Hence the results support the working hypothesis that the dinoflagellates tend to escape the hosts when exposed to stressor-like high temperatures. If the effects were bleaching of the dinoflagellates then we would have seen lower chlorophyll in higher number of zooxanthellae at higher temperature regimes compared to control as growth advanced. Further, there should have been a temperature-dependent decrease in chlorophyll content in the dinoflagellates, which is clearly not the case.
Conclusion
Notwithstanding, it would be desirable to repeat the experiments with more animals to get statistically relevant results. Moreover, the animals from fields should be first acclimatized to the lab conditions for at least two generations under control conditions before the experiment on high temperature regime is conducted. With these precautions the hypothesis is expected to be more rigorously tested. To conclude, within the experimental limitations, a reduction in zooxanthellae population seems to be the primary cause of Cnidarian bleaching.
References
- Clayton, W.S. & Laskerh, H.R.(1984). Host Feeding Regime and Zooxanthellal Photosynthesis in the Anemone, Aiptasia Pallida (Verrill). Biological Bulletin, 167, 590-600.
- Ralph, P.J., Gademann, R. & Larkum, A.W.D.(2001). Zooxanthellae expelled from bleached corals at 33oC are photosynthetically competent. Marine Ecology Progress Series, 220, 163-168.
- Sawyer, S.J. & Muscatine, L.(2001). Cellular mechanisms underlying temperature- induced bleaching in the tropical sea anemone Aiptasia pulchella. The Journal of Experimental Biology, 204, 3443-3456.
- Weis, V.M.(2008). Cellular mechanisms of Cnidarian bleaching: stress causes the collapse of symbiosis. The Journal of Experimental Biology, 211, 3059-3066.